Synthetic Cell Eats, Grows, Divides
SpudCell could become the hydrogen atom for synthetic biology.

I.
Humans have long wanted to create life. This seems to be an eternal pursuit, driven by some perpetual brainworm nestled deeply in the human mind. And when I say “create life,” I don’t mean to simply have sex and then, nine months later, watch as a baby emerges into this world, kicking and thrashing. I mean instead to literally build an organism piece by piece, molecule by molecule, until inert ingredients give rise to life. Such a feat would demonstrate total mastery over nature and refute all the haters who say life is “inscrutable,” “irreducible,” or “hopelessly” complex.
Alas, no one has yet created a living cell from purified molecules, and probably won’t for many years to come. But scientists have tried!
Most claims of success, though, come from hype-men who cut corners and don’t actually know the raw ingredients of their organisms. Anybody can take an existing lifeform, cut out some pieces, and then claim to have made something new; or, rather, to suck the cytoplasm and nucleus out of one cell, inject them into another, and then claim to have made a “chimeric” lifeform.1
(Even credible claims are often misinterpreted by a sensationalist press. When German-American biologist Jacques Loeb used salts to coax a sea urchin egg to divide, the Boston Herald headline declared, “Creation of Life… Lower Animals Produced by Chemical Means.” The subheading was even better: “Immaculate Conception Explained.”)2
But in a preprint released today, scientists claim they’ve made the first cell capable of “feeding, growth, replication, division and selection as a single cycle, coupled to one genome, entirely using components scientists put there.” The cell is called SpudCell, and it is made entirely from known molecules. And although it is definitely a cell (in that it has a membrane with molecules inside) it is definitely not alive, because it cannot grow indefinitely, it cannot survive without human help, it cannot make its own ribosomes, it cannot break down or recycle waste, and it “dies” after a few divisions.
Despite its wimpiness (their term, not mine), SpudCell will be a useful model for future research. Now that researchers have proven they can build a primitive cell capable of division and growth from simple molecules, others can build upon their work.
One reason that biology is so difficult to engineer is because it is infinitely convoluted. Every organism has billions of years of evolutionary baggage, which means every organism behaves a little bit uniquely, which means a discovery made in E. coli doesn’t always work in Bacillus subtilis, and vice versa. (“The only rule in biology is that there are exceptions to every rule.”) Each biologist studies their own organism, with its own little quirks, using slightly bespoke tools. This makes comparisons between laboratories frustratingly difficult, and means that progress is far slower than it ought to be.
Contrast this with physics, where scientists have performed thousands of experiments using hydrogen, a humble atom with one proton and one electron. During the 20th century, it was hydrogen that enabled physics to develop a quantum theory of matter, simply because hundreds of scientists were using precisely the same type of atom for their experiments. Scientists could directly compare, criticize, and argue over their results, because everybody was working in the same medium.
Biology has no hydrogen atom. E. coli may be the most well-studied organism of all time, but hundreds of its genes still have unknown functions. So perhaps SpudCell will become that hydrogen atom; the model that finally enables biologists to fully dissect the mechanisms of life.
II.
Synthetic biologists have been rebuilding parts of cells from the bottom up for decades, or reconstituting cellular functions using purified components. It’s just that nobody had yet managed to put these pieces together into something that behaves — even a little bit — like a living cell.
In 2001, a Japanese group purified dozens of proteins from living cells, recombined them in a tube, and showed that this mixture was sufficient for protein synthesis, meaning the purified molecules could assemble proteins from genetic code. Other groups have reconstituted feeding in test tubes. In 2004, for example, researchers built a small liposome (or bubble enclosed by a lipid membrane), studded its membrane with protein pores, and showed that this liposome could import nutrients. Scientists have even reconstituted cell division! In 2020, a German team stuck bulky proteins onto a lipid membrane and found that those proteins repulsed each other, bent the membrane and, eventually, coaxed the cell to split.3
But these are isolated examples. None of these reconstituted mixtures could eat, grow, or express genes at the same time. The more promising route toward synthetic life, then, has been to work from the top-down. Instead of assembling a cell from chemicals, researchers can instead take an existing organism and ruthlessly cut out its DNA, gene by gene, until arriving at the minimal set required for life.
In 2016, for example, the J. Craig Venter Institute released JCVI-syn3.0, a Mycoplasma mycoides cell it had reduced to 473 genes and 531,000 bases of DNA (far smaller than any self-replicating cell found in nature. Its natural genome has about 1.2 million basepairs of DNA.)4 About 149 of those genes don’t have a known function, but the cell has a metabolism, disposes of waste, and keeps dividing indefinitely; about once every three hours.
These two approaches — bottom-up and top-down — are complementary. Stripping an organism down to only the genes it requires, and then working out what each one does, is a perfectly valid way to build a simplified lifeform. But the genes that remain, after all that chopping, are what’s necessary for the cell to divide, and necessity is not the same as understanding. Kate Adamala, corresponding author of the SpudCell paper, argues that the only way to truly understand how life works, at a molecular resolution, is to assemble it from scratch.
“If we lose track of what’s in [the cell], then what’s the point?” she says. “The whole point of engineering a synthetic cell, for me, is that I need to know all the molecules.”
III.
SpudCell is a lipid blob filled with proteins and DNA. It’s probably easiest to understand how it works by looking at each “module” independently.
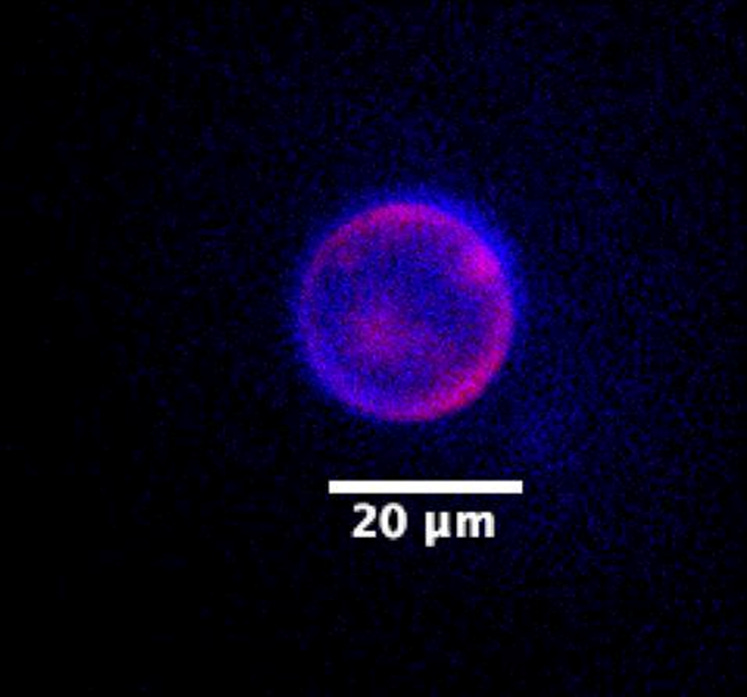
Let’s start with the genome, which spans seven plasmids, or loops, built from 90,000 total base pairs of DNA. Each plasmid encodes one or more genes. One plasmid encodes T7 RNA polymerase, an enzyme that transcribes DNA into RNA; another encodes Phi29, an enzyme that copies DNA; and other plasmids encode α-hemolysin (more on this later), green fluorescent protein, and multiple “accessory” proteins that help with protein synthesis. Each plasmid also has a promoter, or short snippet of DNA, that signals RNA polymerase to switch on the gene.
DNA replication is simple; the Phi29 protein diffuses around SpudCell, randomly grabs onto plasmids, and then copies them. This happens continuously, and the energy and nucleotides needed for DNA replication all come from outside, not from the cell itself. Transcription, or converting DNA into RNA, is also simple. The T7 RNA polymerase floats around, binds to promoters, and turns each gene into messenger RNA.
Protein synthesis is more complicated, because dozens of proteins (and RNAs) are involved. There is the ribosome, of course, but also tRNAs, enzymes that make amino acids, and more.5 SpudCell does not genetically encode some of these other molecules, including the ribosomes. Instead, the researchers pack them into the lipid bubble at the start of the experiment and then, to keep the cell growing, supply additional ribosomes as food.
But SpudCell doesn’t eat food like most living organisms do. Whereas a normal cell pulls in nutrients through protein channels embedded in its membrane, SpudCell instead feeds by merging its body with lipid bubbles.
Remember the α-hemolysin protein I mentioned earlier? Well, that’s what SpudCell uses to eat. Specifically, the researchers engineered the α-hemolysin proteins to carry histidine tags, or short strings of histidine amino acids, on the portion that sticks out from the cell. These tags latch onto feeder liposomes — basically lipid bubbles filled with energy, nutrients, and ribosomes — and coax them to merge into SpudCell and dump that food inside.6
The authors argue that this is a “genetically encoded form of feeding,” because one of the proteins encoded on the plasmids (α-hemolysin) mediates it. Which, sure, that is technically true. But in reality, SpudCell has no metabolism. It can’t break down waste, recycle nutrients, or reuse atoms, which means it can’t survive for long (a problem I’ll return to later).
And what about cell division? That’s probably the hardest thing to reconstitute from the bottom up, because it requires so many steps: SpudCell must copy its plasmids, partition those copies to each half of the cell, and then pinch the membrane in two. The cytoskeleton does this in normal cells, but SpudCell has no such cytoskeleton, so it instead divides in a much cruder way, using the same α-hemolysin protein that feeds it.
The trick relies on a beautiful idea from membrane biophysics. If you stud the outside of a lipid membrane with bulky proteins, even at a low density, then the membrane will start to bend. The bulky proteins crowd and repel one another, thus forcing the membrane to curve away. As more and more proteins pack the membrane, this strain eventually forces it to split.7
SpudCell does much the same, again using those histidine tags on α-hemolysin. But this time, instead of grabbing a feeder liposome, each tag grabs onto a small linker molecule that the researchers supply into the surrounding environment. One end of that linker binds the histidine tag; the other end carries biotin. And biotin, in turn, binds tightly to streptavidin — the bulky protein that actually does the work (and which is supplied by the researchers along with biotin). As more and more streptavidin proteins grab onto the α-hemolysin proteins, the more they crowd the SpudCell surface. The membrane bends further and further, until finally the cell pinches in two.
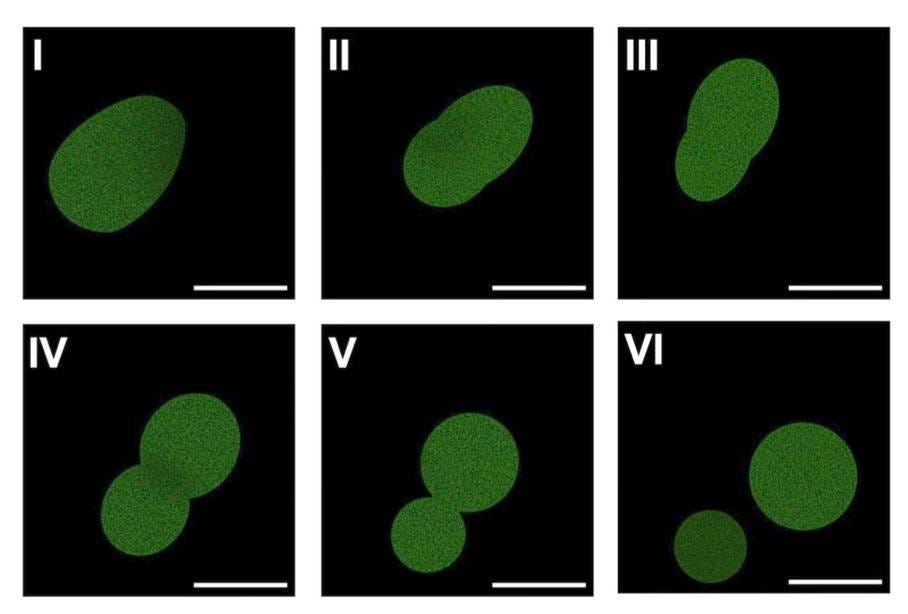
But this doesn’t work reliably! SpudCell only divides a couple of times in this way before dying out. “It’s not a very good system,” says Adamala. “It’s wimpy.”
Wimpiness aside, the fact that α-hemolysin — a protein encoded by the SpudCell genome — drives both feeding and cell division means that SpudCell is subject to selection, whether natural or artificial.
To prove this, the researchers mutated the promoter next to the α-hemolysin gene to make it “stronger,” or more likely to attract RNA polymerase. Cells with this mutation made more α-hemolysin mRNA, and thus more protein, and thus were more likely than cells without the mutation to consume food and recruit streptavidin for cell division.
When researchers mixed the mutated SpudCells with their normal counterparts at a 50:50 ratio, and then ran them through five generations of growth and division, they found that the mutated variants outcompeted the others and grew to about 60 percent of the total population.
This is artificial selection, and not open-ended evolution. But simplicity is the whole point of this paper! At every step, the researchers chose to implement a simplified biochemical system rather than mimic real cells. Their goal was to get something that works first, and then find ways to make it better over time. But with that in mind, it’s worth thinking about what needs to happen next to make SpudCell capable of long-term growth and division.
One issue is the DNA, which is currently split across seven plasmids. There are many valid reasons to split the genome in this way; it reduces crosstalk between genes, for example, and makes it easier to debug genes when they don’t work. Phi29, the DNA copier, also works best on small pieces of DNA and might struggle to replicate a single 90-kilobase genome.
But because SpudCell doesn’t have a cytoskeleton, it partitions its plasmids randomly during each division. In other words, each plasmid ends up in a random daughter cell, and there is always a chance that one of the daughters won’t get all seven plasmids. Without all the plasmids, the cell will “die.”
(Say the cell has five copies of each of the seven plasmids. In that scenario, there is only an 80 percent chance that a daughter cell will get at least one copy of each plasmid. After five rounds of cell division, only 30 percent of cells will have all seven plasmids.)
Metabolism, though, is probably the biggest issue for long-term growth. Remember that SpudCell can’t make its own ribosomes or energy, so the researchers pump in fresh supplies constantly. This is fine, in principle, but SpudCell also can’t recycle its broken proteins. Ribosomes wear out and proteins have half-lives. When these proteins “break,” they simply sit there and fill up space.8 With each cell division, a larger and larger fraction of the cell’s internal volume becomes packed with junk. The next step to make SpudCell grow for longer, then, is probably to find a way to recycle or remove broken proteins.
This is particularly important for α-hemolysin. Remember that these proteins sit in the cell membrane, where their histidine tags link up with either feeder liposomes or streptavidin. This linkage to streptavidin, in particular, is really difficult to undo! The used up α-hemolysin proteins just sit in the membrane and take up space, then, and as SpudCell divides, this useless junk studs more and more of its membrane.
If the researchers figure out how to solve these problems and stretch the division cycles to 20 or 30 generations, far more interesting experiments will become possible. For example, what if you could swap in an error-prone DNA polymerase — an enzyme that randomly introduces mutations — and then follow the mutating cells over multiple generations to see if they evolve new functions? You could even feed the SpudCells random genes, coax them to incorporate that DNA, and see whether they repurpose it for new functions.
IV.
With all these caveats, and all the comments on SpudCell being “wimpy,” it’s worth thinking about why this paper matters at all.
One reason is that this is the first time anyone has combined all the in vitro modules into a single system that works. SpudCell only sortaworks, and the authors admit as much, but that’s okay; at least biologists now have a platform to add new modules, like a metabolism or a cytoskeleton, and see whether they work as part of a complete system.
For this reason, SpudCell really does feel like it could become the hydrogen atom for biology. And to make that vision a reality, Kate Adamala (together with Drew Endy, Jan Jedryszek, and Chris Raggio) are launching a nonprofit organization, called Biotic, to turn bottom-up cell assembly into an engineering discipline. Biotic has already raised more than $6 million in philanthropic funding.
“The main goal is to unify [the field],” says Adamala. “People make awesome modules, but you try to combine two different modules and you pretty much have to re-engineer half of it from scratch, because it wasn’t made to operate on the same chassis. It’s kind of like writing apps for an operating system. You’ve got to declare which operating system your app is going to work on.”
Biotic will release step-by-step protocols for building cells, open-source its plasmids and components, and fund research that directly improves SpudCell. For example, Adamala plans to consolidate the seven plasmids into a single chromosome, solve the partitioning problems, and engineer these cells to make their own ribosomes from scratch.
Progress, one hopes, will be swift. Biology has too many unknowns; most experiments don’t replicate; and few people truly work on the exact same organism using reliable, replicable methods. If we’re ever going to understand an organism completely, we may need to build it from the bottom up, using solely molecules we put there by design.
Thanks to Richard Murray for helpful comments. Mistakes are my own.
In 1970, biologists in New York published a provocative paper titled “Reassembly of Living Cells from Dissociated Components.” They basically sucked the nuclei out of Amoeba proteus cells, used a micro-needle to extract cytoplasm (about 75 percent of the total volume), and then injected both cytoplasm and a nucleus from another Amoebaback into the dead cells. The “reassembled” Amoebasurvived about 80 percent of the time.
Even presidents fall victim to the hype. When Arthur Kornberg made some viral DNA and showed it was infective, Lyndon Johnson told an audience, “Some geniuses at Stanford University have created life in the test tube!”
The bulky proteins repulse each other, forcing the membrane to bend away from them.
Its natural genome has about 1.2 million basepairs of DNA.
Ribosomes are more than half RNA by mass; so they are really less protein and more ribonucleotide machine!
The feeder liposomes are studded with a molecule called nickel-nitrilotriacetic acid, which latches onto the histidine tags.
The approach was inspired by work from Reinhard Lipowsky, a German physicist. As Adamala puts it, “if you stuff giant GFP proteins onto the outside of a liposome, the membrane starts bending, and eventually dividing.”
The α-hemolysin pores can export some small molecules, but not large proteins.